Implementing effective vaccination plans: overview of immunological considerations
The primary aim of vaccination is to induce immunological memory to specific pathogens without causing disease, writes Tom McNeilly BSc BVM&S PhD MRCVS, head of disease control at Moredun Research Institute
On encountering the pathogen, a specific immune response is induced that clears the infection before the pathogen can cause disease. The specificity of the immune response is due to the expansion of B and T lymphocytes, which specifically recognise components (antigens) of the pathogen through B cell receptors (BCR) and T cell receptors (TCR) on their cell surface. B cells then produce antibodies specific for the pathogen, which can block pathogen entry or aid pathogen clearance, whereas T cells either coordinate the immune response (CD4+ helper T cells) or specifically recognise and kill infected cells (CD8+ cytotoxic T cells). For vaccines to provide optimal protection, they need to do the following:
- Protect animals at an age when the pathogen is most likely to cause disease;
Generate an immune response at the site of infection; and - Generate the most appropriate immune response (for example, antibodies and cytotoxic T cells) for protection.
As bovine respiratory disease (BRD) viruses initiate infection within the respiratory tract mucosa, optimal vaccine-induced protection should involve the induction of respiratory mucosal immune responses, including mucosal antibodies to block virus entry into respiratory epithelial cells and cytotoxic T cells to target and kill virus-infected cells.
However, induction of mucosal responses through vaccination remains challenging, as vaccines delivered via conventional systemic routes (for example, IM or SC) are, generally, poor inducers of mucosal immune responses. To date, only vaccines delivered directly on to mucosal surfaces are consistently effective at inducing mucosal immune responses – the so-called 'mucosal vaccination'. However, mucosal vaccination remains a significant challenge, as such vaccines need to overcome a formidable array of host defences – including dilution and enzymatic degradation in mucosal secretions and exclusion from the host tissue by the mucus layer and the epithelial barrier – before they can induce an immune response. Furthermore, vaccines delivered at mucosal surfaces also need to trigger innate (non-specific) immune responses to ensure the immune system reacts to, rather than 'tolerates', the vaccine. Mucosal vaccines based on live attenuated pathogens, rather than killed pathogens or pathogen fractions, are generally most effective at inducing mucosal immune responses, as they can more effectively overcome the mucosal barriers, efficiently stimulate innate immune responses and, as they closely mimic natural infections, elicit the most appropriate adaptive (specific) immune responses for clearance of the target pathogen. The following sections will provide a brief overview of how immune responses to BRD vaccines are specifically induced at mucosal surfaces, including why delivery of the vaccine to the respiratory tract is important, the role of innate (non-specific) and adaptive (specific) immune responses in controlling BRD viral infections and the influence of maternal antibodies on vaccine-mediated protection.
How mucosal immune responses are induced
As previously stated, the most effective way of inducing mucosal immune responses is by administration of vaccines on to the mucosal surface (for example, intranasal vaccination in the case of BRD viral vaccines, Figure 1). Importantly, while B cells can recognise antigens directly (that is, they can bind parts [epitopes] of a molecule directly through their BCR),
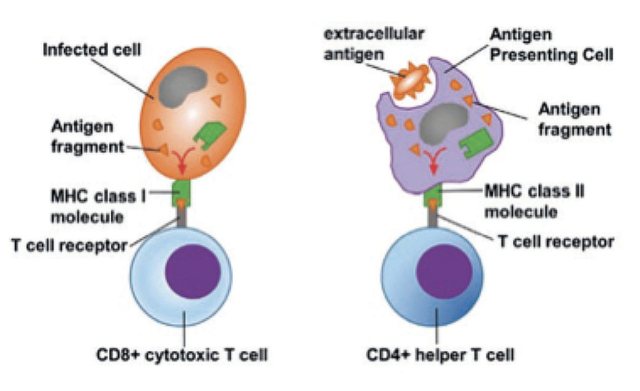
T cells are only able to recognise protein antigens once they have been taken up by antigen-presenting cells (APC), processed into small fragment peptides and then complexed with major histocompatibility complex (MHC) molecules on the APC surface. Furthermore, CD4 T cells only recognise peptides presented by MHC class II molecules, which generally represent extracellular antigens taken up by the APC (endogenous antigen). In contrast, CD8 T cells only recognise antigens presented by MHC class I molecules, which represent intracellular antigens synthesised within the presenting cell itself (endogenous antigens) or extracellular antigens that are taken up by specialised APC called dendritic cells (DC) and then 'cross-presented' on to MHC class I molecules (Figure 2). This makes CD8 T cells excellent at detecting virus-infected cells, as viral proteins are made within host cells and will be presented by MHC class I molecules.
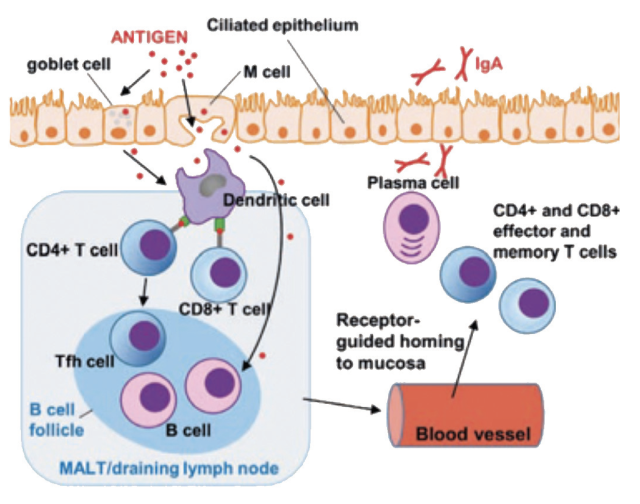
Following mucosal delivery of a vaccine, antigens are primarily sampled by specialised cells (M-cells or goblet cells), which deliver antigen to underlying DC. DCs subsequently process and present antigen to T cells via MHC I and II molecules within collections of lymphoid tissue within the mucosa, known as mucosa-associated lymphoid tissue, or within mucosal lymph nodes to initiate the adaptive immune response1. Soluble or cellbound antigen also traffics directly to these lymphoid tissues to initiate B cell responses2. Specific immune-signalling events, largely mediated by cytokines, during this initiation of the immune responses play a critical role in determining the balance of cellular immune responses versus antibody responses – the so-called T helper type 1 (Th1) and Th2 responses, respectively. During the process of antigen recognition by T and B lymphocytes at the mucosa, expression of mucosal tissue-specific adhesion molecules and chemokine receptors are induced on activated T and B cells, which allow lymphocytes to home back to mucosal sites through recognition of counter receptors expressed on mucosal blood vessels and tissues3. This then results in production of immune effectors (mucosal antibodies, cytotoxic T cells) at the mucosal site. The reason why vaccines delivered via SC or IM routes are poor at inducing mucosal immune responses is that they do not generally confer mucosal homing on lymphocytes. A schematic representation of the induction of mucosal immune responses is shown in Figure 3.
Role of innate immunity in protection against BRD viruses
Following infection of the respiratory tract, BRD viruses initially encounter and infect the respiratory epithelia4,5. This infection rapidly induces expression of a range of pro-inflammatory cytokines and chemokines, which result in the recruitment of innate immune cells, such as neutrophils, to the site of infection6. This innate immune response can be beneficial: for example, some cytokines – such as type I interferons released during inflammation – have antiviral effects on neighbouring uninfected cells, and the innate immune response will direct the subsequent adaptive immune response to ensure it is most capable of clearing the infection (for example, by enhancing CD8+ cytotoxic T cell responses to kill infected cells). However, if the response is uncontrolled, the innate immune response will cause disease. One benefit of live attenuated viral vaccines delivered mucosally is that, unlike natural viral infections, components of the innate immune response will be elicited that will provide some degree of protection within a few days of vaccination without causing disease.
Role of adaptive immune responses: antibody protection versus cell-mediated immunity
The two major effectors of the adaptive immune response are production of antigen-specific antibodies and CD8+ cytotoxic T cells, and both are thought to play a role in protection against BRD viruses. With regard to the antibody response, it appears levels of virus-specific IgA within mucosal secretions are most associated with protection7, whereas elegant studies in which different types of T cells were depleted from calves challenged with bovine respiratory syncytial virus (BRSV) – one of the most important BRD viruses – clearly show CD8+ cytotoxic T cells play a dominant role in the elimination of virus from both the upper and lower respiratory tract.8,9 Importantly, live attenuated BR SV vaccines are better at priming for lung CD8+ T cell responses compared to killed vaccines.10
Impact of maternal antibodies on BRD vaccines
As severe BRD viral infections tend to occur in young calves (usually between one and three months11), vaccines should ideally work in the face of maternally derived antibodies (MDA) and be induced rapidly. However, MDA can interfere with the response to BRD virus vaccination if the antibodies bind to the virus targeted by the vaccine. Interestingly, while antibody responses to the vaccines are suppressed, priming of T cell responses is largely unaffected. Two major mechanisms exist by which MDA is thought to interfere with the generation of antibody responses to vaccines.12 In the first, 'epitope masking', maternal antibodies bind to the vaccine antigens and prevent recognition of the antigens by B cells by interfering with BCR-antigen binding. In the second mechanism, maternal antibodies can form complexes with the vaccine antigens that are then able to cross-link BCRs (which recognises the vaccine antigen) with FcγRIIB, an inhibitory receptor (which recognises the non-antigen binding region of the antibody) on the surface of B cells. This, in turn, inhibits B cell activation and prevents generation of a specific antibody response.
The impact of MDA on vaccine performance largely depends on the mechanism by which the vaccine induces protection. If T cells play a major role in protection, the influence of maternal antibodies will be less and protection in the face of MDA will be achievable as T cell responses are largely unaffected; however, if antibodies are important, then maternal antibodies will have more of an impact. As T cells (specifically CD8+ cytotoxic T cells) play a major role in protection against BRD viruses, early immunisation should afford protection.
However, one disadvantage to vaccination in the face of MDA is that induction of neutralising antibodies that block viral infection of cells will be compromised. Good evidence shows MDA interference with the antibody response to BRD virus vaccines13-15. However, despite this, multiple studies have shown vaccine protection against virus challenge can be achieved in young calves from three days of age13,14,16, and live attenuated viral vaccines delivered mucosally (intranasally) are more effective at inducing protection in the face of MDA than inactivated or live vaccines delivered by injection17. This is presumably due to the induction of innate immune responses and effective priming of cellular immune responses within the respiratory tract. It may also reflect lower levels of virus-specific maternal antibodies at the mucosal surface compared to SC or IM injection sites, and, therefore, less MDA interference with the mucosally delivered vaccines. However, while intranasal vaccination of young calves is achievable, the duration of immunity appears to be short (less than four months), and repeat vaccination may be required to extend the duration of protection.14,18
Conclusions
Effective BRD virus vaccines need to generate effective immune responses within the respiratory tract at an early age. The most efficient way to do this is through live attenuated viral vaccines delivered on to the respiratory tract: these vaccines induce both innate and adaptive immune responses that contribute to the protection against BRD viruses, and, importantly, work in the face of MDA, allowing their use in young calves.
This article was first published in Vet Times, Livestock Supplement.
Biography
Tom McNeilly is head of disease control at Moredun Research Institute. He is a qualified veterinary surgeon, and spent four years in mixed practice before embarking on a career in research in 2006. Tom’s major research interests are in mucosal immunology, specifically host-pathogen interactions at mucosal surfaces, and the development of vaccines to control endemic livestock diseases. Mr McNeilly wrote this article on behalf of Boehringer Ingelheim Animal Health.
- Neutra MR et al (2001). Collaboration of epithelial cells with organized mucosal lymphoid tissues, Nature Immunology 2(11): 1,004-1,009.
- Harwood NE and Batista FD (2010). Antigen presentation to B cells, F1000 Biology Reports 2: 87.
- Kunkel EJ and Butcher EC (2003). Plasma-cell homing, Nature Reviews: Immunology 3(10): 822-829.
- Kirchhoff J et al (2014). Three viruses of the bovine respiratory disease complex apply different strategies to initiate infection, Veterinary Research 45(1): 20.
- Cusack PM et al (2003). The medicine and epidemiology of bovine respiratory disease in feedlots, Australian Veterinary Journal 81(8): 480-487.
- Gershwin LJ (2012). Immunology of bovine respiratory syncytial virus infection of cattle, Comparative Immunology, Microbiology and Infectious Diseases 35(3): 253-257.
- Kimman TG et al (1987). Local and systemic antibody response to bovine respiratory syncytial virus infection and reinfection in calves with and without maternal antibodies, Journal of Clinical Microbiology 25(6): 1,097-1,106.
- Taylor G et al (1995). Role of T-lymphocyte subsets in recovery from respiratory syncytial virus infection in calves, Journal of Virology 69(11): 6,658-6,664.
- Thomas LH (1996). Influence of selective T-lymphocyte depletion on the lung pathology of gnotobiotic calves and the distribution of different T-lymphocyte subsets following challenge with bovine respiratory syncytial virus, Research in Veterinary Science 61(1): 38-44.
- Antonis AF et al (2006). Kinetics of antiviral CD8 T cell responses during primary and post-vaccination secondary bovine respiratory syncytial virus infection, Vaccine 24(10): 1,551-1,561.
- Kimman TG et al (1988). Epidemiological study of bovine respiratory syncytial virus infections in calves: influence of maternal antibodies on the outcome of disease, Veterinary Record 123(4): 104-109.
- Niewiesk S (2014). Maternal antibodies: clinical significance, mechanism of interference with immune responses, and possible vaccination strategies, Frontiers in Immunology 5: 446.
- Ellis JA et al (2010). Response to experimentally induced infection with bovine respiratory syncytial virus following intranasal vaccination of seropositive and seronegative calves, Journal of the American Veterinary Medical Association 236(9): 991-999.
- Ellis JA et al (2013). Duration of immunity to experimental infection with bovine respiratory syncytial virus following intranasal vaccination of young passively immune calves, Journal of the American Veterinary Medical Association 243(11): 1,602-1,608.
- Ellis J et al (2014). Inhibition of priming for bovine respiratory syncytial virus-specific protective immune responses following parenteral vaccination of passively immune calves, The Canadian Veterinary Journal 55(12) 1,180-1,185.
- Xue W et al (2010). Immunogenicity of a modified-live virus vaccine against bovine viral diarrhea virus types 1 and 2, infectious bovine rhinotracheitis virus, bovine parainfluenza-3 virus, and bovine respiratory syncytial virus when administered intranasally in young calves, Vaccine 28(22): 3,784-3,792
- Kimman TG et al (1989). Priming for local and systemic antibody memory responses to bovine respiratory syncytial virus: effect of amount of virus, virus replication, route of administration and maternal antibodies, Veterinary Immunology and Immunopathology 22(2): 145-160.
- Ellis J et al (2018). Comparative efficacy of modified-live and inactivated vaccines in boosting responses to bovine respiratory syncytial virus following neonatal mucosal priming of beef calves, The Canadian Veterinary Journal 59(12): 1,311-1,319.